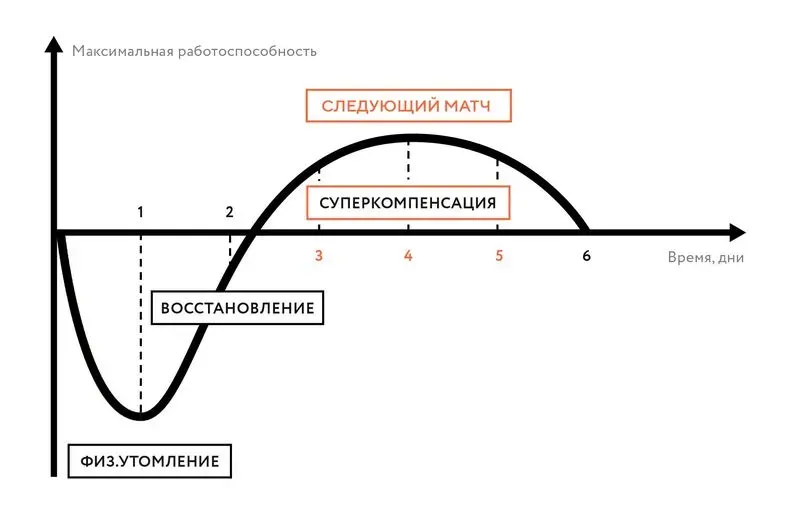
Какие виды мышечных сокращений вы знаете. Режимы сокращения и разновидности работы мышц
 Мышцы (от слова «мышь» - из-за формы, поэтому ударение на первый слог) или мускулы (от лат. musculus - мышца (mus - мышка, маленькая мышь)) - органы тела животных и человека, состоящие из упругой, эластичноймышечной ткани, способной сокращаться под влиянием нервных импульсов. Предназначены для выполнения различных действий: движения тела, сокращенияголосовых связок, дыхания.
Мышцы (от слова «мышь» - из-за формы, поэтому ударение на первый слог) или мускулы (от лат. musculus - мышца (mus - мышка, маленькая мышь)) - органы тела животных и человека, состоящие из упругой, эластичноймышечной ткани, способной сокращаться под влиянием нервных импульсов. Предназначены для выполнения различных действий: движения тела, сокращенияголосовых связок, дыхания.Мышцы позволяют двигать частями тела и выражать в действиях мысли и чувства. Человек выполняет любые движения - от таких простейших, как моргание или улыбка, до тонких и энергичных, какие мы наблюдаем у ювелиров или спортсменов - благодаря способности мышечных тканей сокращаться. От исправной работы мышц, состоящих из трёх основных групп, зависит не только подвижность организма, но и функционирование всех физиологических процессов. А работой всех мышечных тканей управляет нервная система, которая обеспечивает их связь с головным и спинным мозгом и регулирует преобразование химической энергии в механическую.
В теле человека 640 мышц (в зависимости от метода подсчёта дифференцированных групп мышц их общее число определяют от 639 до 850). Самые маленькие прикреплены к мельчайшим косточкам, расположенным в ухе. Самые крупные - большиеягодичные мышцы, они приводят в движение ноги. Самыесильные мышцы- икроножные и жевательные, язык.
По форме мышцы очень разнообразны. Чаще всего встречаются веретенообразные мышцы, характерные для конечностей, иширокие мышцы- они образуют стенки туловища. Если у мышц общее сухожилие, а головок две или больше, то их называют двух-, трёх- или четырёхглавыми.
Мышцы и скелет определяют формучеловеческого тела. Активныйобраз жизни, сбалансированное питание и занятие спортом способствуют развитию мышц и уменьшению объёма жировой ткани.
Режимы сокращения мышц
Для скелетной мышцы характерны два основных режимасокращения - изометрический иизотонический. Изометрический режим проявляется в том, что в мышце во время ееактивности нарастает напряжение (генерируется сила), но из-за того, что обаконца мышцы фиксированы (например, мышца пытается поднять большой груз) - она не укорачивается. Изотонический режимпроявляется в том, что мышца первоначально развивает напряжение (силу),способную поднять данный груз, а потом мышца укорачивается - меняет свою длину, сохраняя напряжение,равное весу поднимаемого груза. Так как изотоническое сокращение не является" чисто" изотоническим (элементы изометрического сокращения имеютместо в самом начале сокращения мышцы), а изометрическое сокращение тоже неявляется " чисто" изотоническим (элементы смещения все-таки есть,несомненно), то предложено употреблять термин " ауксотоническое сокращение"- смешанное по характеру.
Понятия " изотонический", " изометрический"важны для анализа сократительной активности изолированных мышц и для пониманиябиомеханики сердца.
Режимы сокращениягладких мышц. Целесообразно выделитьизометрический и изотонический режимы (и, как промежуточный - ауксотонический). Например, когда мышечнаястенка полого органа начинает сокращаться, а орган содержит жидкость, выход длякоторой перекрыт сфинктером, то возникает ситуация изометрического режима: давлениевнутри полого органа растет, а размеры ГМК не меняются (жидкость не сжимается).Если это давление станет высоким и приведет к открытию сфинктера, то ГМКпереходит в изотонический режим функционирования- происходит изгнание жидкости, т.е. размеры ГМК уменьшаются, анапряжение или сила - сохраняетсяпостоянной и достаточной для изгнания жидкости.
Виды сокращенийУ скелетной мышцы выделяют одиночное сокращение исуммированное сокращение, или тетанус. Одиночное сокращение - это сокращение, которое возникает наодиночный стимул, достаточный для вызова возбуждения мышцы. После короткогоскрытого периода (латентный период) начинается процесс сокращения. Прирегистрации сократительной активности в изометрических условиях (два концанеподвижно закреплены) в первую фазу происходит нарастание напряжения (силы), аво вторую - ее падение до исходнойвеличины. Соответственно эти фазы называют фазой напряжения и фазойрасслабления. При регистрации сократительной активности в изотоническом режиме(например, в условиях обычной миографической записи) эти фазы будут называтьсясоответственно фазой укорочения и фазой удлинения. В среднем сократительныйцикл длится около 200 мс (мышцы лягушки) или 30-80 мс (у теплокровных). Если на мышцудействует серия прямых раздражении (минуя нерв) или непрямых раздражении (черезнерв), но с большим интервалом, при котором всякое следующее раздражениепопадает в период после окончания 2-й фазы, то мышца будет на каждый из этихраздражителей отвечать одиночным сокращением.
Суммированные сокращения возникают в том случае, если намышцу наносятся 2 и более раздражения,причем всякое последующее раздражение (после предыдущего) наносится либо вовремя 2-й фазы (расслабления или удлинения), либо во время 1-й фазы (укороченияили напряжения).
Одиночное сокращение: А - потенциал действия; Б - сокращение мышцы; 1 - фаза напряжения; 2 - фаза расслабления
Суммированное сокращение: а - одиночное сокращение; б-г - зубчатый тетанус; д - гладкий тетанус
В случае, когда всякое второе раздражение попадает в периодфазы расслабления (удлинения), возникает частичная суммация - сокращение еще полностью не закончилось, ауже возникло новое. Если подается много раздражителей с подобным интервалом, товозникает явление зубчатого тетануса. Если раздражители наносятся с меньшиминтервалом и каждое последующее раздражение попадает в фазу укорочения, товозникает так называемый гладкий тетанус.
СТРОЕНИЕ СКЕЛЕТНЫХ МЫШЦ
Скелетные мышцы покрыты толстой соединительнотканной оболочкой. В толще мышцы более тонкая оболочка из соединительной ткани окружает как пучки мышечных волокон, так и отдельные волокна. Эта оболочка выполняет две функции. Во-первых, через неё к мышечным волокнам проходят нервы, кровеносные и лимфатические сосуды. Во-вторых, эластичная соединительная ткань объединяет тянущие усилия множества мышечных волокон.
Мышечные клетки, или волокна, имеют форму заострённых на концах цилиндров. Их тонкая цитоплазматическая мембрана называется сарколеммой, а их цитоплазма - саркоплазмой. По периферии саркоплазмы располагаются многочисленные ядра мышечного волокна, а в центральной части плотно уложены ориентированные продольно миофибриллы; в одном мышечном волокне их может содержаться больше тысячи.
Именно миофибриллы создают картину поперечной исчерченности мышечного волокна, в котором чередуются тёмные и светлые диски. Тёмные диски характеризуются двойным лучепреломлением в поляризованном свете, поэтому их называют анизотропными, в отличие от светлых изотропных дисков (рис. 5). В середине каждого изотропного диска видна тёмная тонкая линия, получившая название Z-мембраны. Участок миофибриллы между соседними Z-мембранами называется саркомером, его длина в среднем составляет около 2-3 мкм.
Саркомеры образованы нитями двух сократительных белков - актина и миозина. Более тонкие нити актина одним концом прикрепляются к Z-мембране. К этой же мембране с другой стороны прикрепляются, как щетинки к щётке, актиновые нити соседнего саркомера. В промежутки между актиновыми нитями примерно на 1/4 их длины входят толстые нити второго сократительного белка - миозина. Участки миофибриллы, содержащие миозин, соответствуют тёмным анизотропным дискам, а актин - светлым изотропным дискам. Мышечное сокращение осуществляется с помощью специального механизма втягивания тонких нитей актина в центр саркомера между толстыми нитями миозина. При сокращении уменьшается только длина саркомеров, тогда как длина актиновых и миозиновых нитей не изменяется.
Каждая тонкая нить актина образована двойной цепью глобулярных молекул мономеров актина, что можно сравнить с двойной ниткой бус, слегка скрученных по оси (рис. 6). На каждой стороне такой двойной цепи есть продольные спиральные желобки. В желобках уложены длинные и тонкие молекулы регуляторного белка тропомиозина. К нитям тропомиозина присоединены молекулы второго регуляторного белка - тропонина, расположенные на одинаковом расстоянии друг от друга. Регуляторные белки контролируют соединения между нитями актина и миозина: от этого зависит сокращение и расслабление мышцы. Например, в расслабленной мышце молекулы тропомиозина занимают положение, препятствующее соединению миозиновых нитей с актиновыми.
У толстых нитей миозина есть многочисленные боковые ответвления, или поперечные мостики, имеющие форму двойных головок (рис.6). Эти головки могут поворачиваться, как на шарнирах, прикрепляться к нитям актина, а затем создавать тянущее усилие. Половина головок каждой нити миозина обращена в одну сторону, а половина - в другую, следовательно каждая половина головок может присоединиться к нитям актина, связанным как с одной, так и с другой Z-мембранами саркомера.
Рис. 6. Сократительные и регуляторные белки миофибрилл. В расслабленной мышце тропомиозин препятствует образованию поперечных мостиков между актином и миозином. При возбуждении мышцы в саркоплазме повышается концентрация ионов кальция, которые соединяются с тропонином, что меняет конформацию его молекул. В результате этого тропонин смещает молекулу тропомио-зина в глубину желобка актиновой нити: тем самым головкам миозина предоставляется возможность соединяться с нитями актина
От сарколеммы мышечного волокна отходят внутрь узкие канальцы, получившие название поперечных Т-трубочек. Когда возбуждение передаётся через нервно-мышечный синапс на сарколемму, волна деполяризации распространяется и на Т-трубочки. В саркоплазме с Т-трубочками соседствуют цистерны саркоплазматического ретикулума, представляющего мышечный аналог гладкого эндоплазматического ретикулума. Вокруг одного саркомера проходят две Т-трубочки, а к каждой из них с обеих сторон прилежат цистерны саркоплазматического ретикулума. Каждая Т-трубочка с двумя прилежащими к ней цистернами саркоплазматического ретикулума образует триаду. Цистерны ретикулума соединены друг с другом посредством множества продольных трубочек, образуя единую систему. В саркоплазматическом ретикулуме накапливаются ионы кальция, поступающие из саркоплазмы активным транспортом. Главный интегральный белок мембраны саркоплазматического ретикулума выполняет функцию кальциевого насоса и при расслаблении мышцы перекачивает ионы кальция в ретикулум, используя для этого энергию АТФ.
НЕЙРО-МОТОРНЫЕ ЕДИНИЦЫ
Структурно-функциональной единицейскелетных мышцявляется многоядерное мышечное волокно. Объединяясь в пучки, эти волокна образуют мышцу. Любое мышечное волокно иннервируется только от одного мотонейрона. В то же время один мотонейрон иннервирует группу мышечных волокон, вместе с которыми он образует двигательную единицу. Она является наименьшей функциональной единицей моторной системы. Вразных мышцахдвигательные единицы сильно отличаются по количеству мышечных волокон. Так, например, в глазо-двигательных мышцах на один нейрон приходится около десятка мышечных волокон, в двигательных единицах мелких мышц руки их около сотни, а в крупных мышцах конечностей и туловища - более тысячи на один мотонейрон. Малые единицы характерны для мышц, с помощью которых совершаются точные, хорошо координированые движения, тогда как большие преобладают в массивных мышцах, таких например, как мышцы спины.
В зависимости от скорости развития максимальной силы сокращения, а также от быстроты утомления различают два типа двигательных единиц: 1) быстрый, утомляемый тип; 2)медленный, мало утомляемый. В большинстве мышц представлены два типа двигательных единиц, но в разном соотношении, которое варьирует у людей индивидуально. В спортивной физиологии людей с относительно большим процентом быстрых волокон относят к спринтерам, а обладателей большего, чем в среднем, процента медленных волокон - к стайерам. У всех людей при старении процент быстрых волокон уменьшается.
Волокна быстрых двигательных единиц относительно меньше снабжаются кровью и поэтому имеют бледно-красный цвет. Они предназначены для мощных быстрых сокращений в течение короткого времени, поскольку быстро утомляются, не поспевая из-за относительно малого кровоснабжения окислять глюкозу до конечных продуктов. Волокна медленного типа обеспечены обильной сетью кровеносных капилляров, что придаёт им тёмно-красный цвет. И, хотя они сокращаются медленнее, зато способны очень долго работать без утомления. Развиваемая ими сила составляет от 1 до 10% от силы быстрых волокон.
Тела мотонейронов двигательных единиц медленного типа имеют наименьшие размеры и самый низкий порог возбуждения, поэтому только они могут активироваться слабыми входными сигналами. Мотонейроны быстрых двигательных единиц, напротив, являются самыми крупными и, чтобы возбудить их, нужны сильные стимулы. Поэтому медленные двигательные единицы используются при любых движениях, а быстрые - только в тех случаях, когда понадобится большая сила сокращающихся мышц. Так, например, при стоянии или спокойной ходьбе сокращение мышц обеспечивает менее чем половина всех двигательных единиц - здесь сила мышечного сокращения составляет около 20% от максимальной. Для того чтобы выполнить прыжок, может потребоваться максимальная сила: в таком случае понадобится активация мотонейронов быстрых двигательных единиц.
ВИДЫ И РЕЖИМЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ.
Мышечное сокращение проявляется в укорочении мышцы. В зависимости от условий стимуляции и функционального состояния мышцы может возникнуть одиночное и тетаническое сокращение мышцы.
Амплитуда одиночного сокращения зависит от количества сократившихся в этот момент миофибрилл. Возбудимость отдельных волокон, составляющих целую мышцу, различна, поэтому пороговая сила тока вызывает сокращение лишь наиболее возбудимых мышечных волокон. Амплитуда такого сокращения минимальна.
Мышечному сокращению предшествует процесс возбуждения (т.е. ПД), который совпадает по времени с латентным периодом мышечного сокращения.
Рис. 7. Соотношения фаз сократимости(А) и возбудимости(Б) поперечно-полосатых мышц:
1,6 - исходная возбудимость; 2,5 - супернормальная возбудимость; 3 - абсолютная рефрактерность; 4 - относительная рефрактерность
Сила и скорость мышечного сокращения пропорциональны также частоте потенциалов действия, распространяющихся к мышце по аксону мотонейрона - это ещё один механизм регуляции, который называется частотным кодированием. Потенциал действия в нерве или мышце длится приблизительно 1-3 мс, а самое короткое сокращение, состоящее из укорочения и расслабления, продолжается примерно 100 мс (рис 7). Таким сокращением мышца отвечает на единичный потенциал действия - это сокращение называется одиночным (рис.8).
Рис.8 - Одиночноемышечное сокращение.
1 - латентная фаза; 2 - фаза сокращения; 3 - фаза расслабления
Если частота возбуждающих мышцу нервных импульсов станет расти, она не успеет полностью расслабиться к моменту прихода очередного потенциала действия, и сила её сокращения будет увеличиваться. Такой вид сокращения получил название зубчатого тетануса. С ещё большим увеличением частоты стимуляции формируется плато: в это время мышца совсем не расслабляется, а сокращение делается максимальным - этот режим называется гладким тетанусом (рис.9).
|
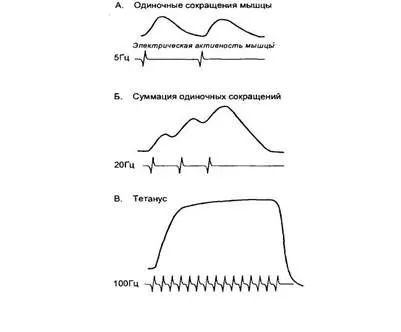
А. При малой частоте нервных импульсов, возбуждающих мышцу, она отвечает на каждый из них одиночным сокращением, успевая расслабиться до прихода следующего импульса;
Б. С увеличением частоты нервных импульсов происходит суммация одиночных сокращений, при которой растёт амплитуда сократительных ответов;
В. При достаточно большой частоте нервных импульсов мышца не может расслабиться, а сократительный ответ становится максимальным.
Серия следующих друг за другом потенциалов действия приводит к значительному повышению внутриклеточной концентрации кальция, поэтому сократительный ответ и становится сильнее и продолжительнее. Частота поступающих к мышцам потенциалов действия варьирует в небольших пределах. Например, при произвольных сокращениях потенциалы действия в мотонейронах появляются с частотой приблизительно 8 Гц, в редких случаях она может превысить 25 Гц. С нарастанием частоты поступающих к мышце потенциалов действия сила её сокращения увеличивается (рис.10).
|
Похожая информация.
Мышечное сокращение является жизненноважной функциейорганизма, связанной с оборонительными, дыхательными, пищевыми, половыми, выделительными и другими физиологическими процессами. Все виды произвольных движений – ходьба, мимика, движенияглазных яблок, глотание, дыхание и т. п. осуществляются за счет скелетных мышц. Непроизвольные движения (кроме сокращения сердца) – перистальтика желудка и кишечника, изменение тонуса кровеносных сосудов, поддержание тонуса мочевого пузыря – обусловлены сокращением гладких мышц. Работа сердца обеспечивается сокращением сердечной мускулатуры.
Структурная организация скелетной мышцы
Мышечное волокно и миофибрилла (рис. 1). Скелетная мышца состоит из множества мышечных волокон, имеющих точки прикрепления к костям и расположенных параллельно друг другу. Каждое мышечное волокно (миоцит) включает множество субъединиц – миофибрилл, которые построены из повторяющихся в продольном направлении блоков (саркомеров). Саркомер является функциональной единицей сократительного аппарата скелетной мышцы. Миофибриллы в мышечном волокне лежат таким образом, что расположение саркомеров в них совпадает. Это создает картину поперечной исчерченности.
Саркомер и филламенты. Саркомеры в миофибрилле отделены друг от друга Z -пластинками, которые содержат белок бета-актинин. В обоих направлениях от Z -пластинки отходят тонкие актиновые филламенты. В промежутках между ними располагаются более толстые миозиновые филламенты.
Актиновый филламент внешне напоминает две нитки бус, закрученные в двойную спираль, где каждая бусина – молекула белка актина. В углублениях актиновых спиралей на равном расстоянии друг от друга лежат молекулы белка тропонина, соединенные с нитевидными молекулами белка тропомиозина.
Миозиновые филламенты образованы повторяющимися молекулами белка миозина. Каждая молекула миозина имеет головку и хвост. Головка миозина может связываться с молекулой актина, образуя так называемый поперечный мостик.
Клеточная мембрана мышечного волокна образует инвагинации ( поперечные трубочки ), которые выполняют функцию проведения возбуждения к мембране саркоплазматического ретикулума. Саркоплазматичекий ретикулум (продольные трубочки) представляет собой внутриклеточную сеть замкнутых трубочек и выполняет функцию депонирования ионов Са++.
Двигательная единица. Функциональной единицей скелетной мышцы является двигательная единица (ДЕ). ДЕ – совокупность мышечных волокон, которые иннервируются отростками одного мотонейрона. Возбуждение и сокращение волокон, входящих в состав одной ДЕ, происходит одновременно (при возбуждении соответствующего мотонейрона). Отдельные ДЕ могут возбуждаться и сокращаться независимо друг от друга.
Молекулярные механизмы сокращения скелетной мышцы
Согласно теории скольжения нитей, мышечное сокращение происходит благодаря скользящему движению актиновых и миозиновых филламентов друг относительно друга. Механизм скольжения нитей включает несколько последовательных событий.
Головки миозина присоединяются к центрам связывания актинового филламента (рис. 2, А).
Взаимодействие миозина с актином приводит к конформационным перестройкам молекулы миозина. Головки приобретают АТФазную активность и поворачиваются на 120 °. За счет поворота головок нити актина и миозина передвигаются на «один шаг» друг относительно друга (рис. 2, Б).
Рассоединение актина и миозина и восстановление конформации головки происходит в результате присоединения к головке миозина молекулы АТФ и ее гидролиза в присутствии Са++ (рис. 2, В).
Цикл «связывание – изменение конформации – рассоединение – восстановление конформации» происходит много раз, в результате чего актиновые и миозиновые филламенты смещаются друг относительно друга, Z -диски саркомеров сближаются и миофибрилла укорачивается (рис. 2, Г).
Сопряжение возбуждения и сокращения в скелетной мышце
В состоянии покоя скольжения нитей в миофибрилле не происходит, так как центры связывания на поверхности актина закрыты молекулами белка тропомиозина (рис. 3, А, Б). Возбуждение (деполяризация) миофибриллы и собственно мышечное сокращение связаны с процессом элетромеханического сопряжения, который включает ряд последовательных событий.
В результате срабатывания нейромышечного синапса на постсинаптической мембране возникает ВПСП, который генерирует развитие потенциала действия в области, окружающей постсинаптическую мембрану.
Возбуждение (потенциал действия) распространяется по мембране миофибриллы и за счет системы поперечных трубочек достигает саркоплазматического ретикулума. Деполяризации мембраны саркоплазматического ретикулума приводит к открытию в ней Са++ -каналов, через которые в саркоплазму выходят ионы Са++ (рис. 3, В).
Ионы Са++ связываются с белком тропонином. Тропонин изменяет свою конформацию и смещает молекулы белка тропомиозина, которые закрывали центры связывания актина (рис. 3, Г).
К открывшимся центрам связывания присоединяются головки миозина, и начинается процесс сокращения (рис. 3, Д).
Для развития указанных процессов требуется некоторый период времени (10–20 мс). Время от момента возбуждения мышечного волокна (мышцы) до начала ее сокращения называют латентным периодом сокращения.
Расслабление скелетной мышцы
Расслабление мышцы вызывается обратным переносом ионов Са++ посредством кальциевого насоса в каналы саркоплазматического ретикулума. По мере удаления Са++ из цитоплазмы открытых центров связывания становится все меньше и в конце концов актиновые и миозиновые филламенты полностью рассоединяются; наступает расслабление мышцы.
Контрактурой называют стойкое длительное сокращение мышцы, сохраняющееся после прекращения действия раздражителя. Кратковременная контрактура может развиваться после тетанического сокращения в результате накопления в саркоплазме большого количества Са++; длительная (иногда необратимая) контрактура может возникать в результате отравления ядами, нарушений метаболизма.
Фазы и режимы сокращения скелетной мышцы
Фазы мышечного сокращения
При раздражении скелетной мышцы одиночным импульсом электрического тока сверхпороговой силы возникает одиночное мышечное сокращение, в котором различают 3 фазы (рис. 4, А):
латентный (скрытый) период сокращения (около 10 мс), во время которого развивается потенциал действия и протекают процессы электромеханического сопряжения; возбудимость мышцы во время одиночного сокращения изменяется в соответствии с фазами потенциала действия;
фаза укорочения (около 50 мс);
фаза расслабления (около 50 мс).
Рис. 4. Характеристика одиночного мышечного сокращения. Происхождение зубчатого и гладкого тетануса. Б – фазы и периоды иышечного сокращения, Б – режимы мышечного сокращения, возникающие при разной частоте стимуляции мышцы. Изменение длины мышцы показано синим цветом, потенциал действия в мышце - красным, возбудиумость мышцы - фиолетовым. |
Режимы мышечного сокращения
В естественных условиях в организме одиночного мышечного сокращения не наблюдается, так как по двигательным нервам, иннервирующим мышцу, идут серии потенциалов действия. В зависимости от частоты приходящих к мышце нервных импульсов мышца может сокращаться в одном из трех режимов (рис. 4, Б).
Одиночные мышечные сокращения возникают при низкой частоте электрических импульсов. Если очередной импульс приходит в мышцу после завершения фазы расслабления, возникает серия последовательных одиночных сокращений.
При более высокой частоте импульсов очередной импульс может совпасть с фазой расслабления предыдущего цикла сокращения. Амплитуда сокращений будет суммироваться, возникнет зубчатый тетанус – длительное сокращение, прерываемое периодами неполного расслабления мышцы.
При дальнейшем увеличении частоты импульсов каждый следующий импульс будет действовать на мышцу во время фазы укорочения, в результате чего возникнет гладкий тетанус – длительное сокращение, не прерываемое периодами расслабления.
Оптимум и пессимум частоты
Амплитуда тетанического сокращения зависит от частоты импульсов, раздражающих мышцу. Оптимумом частоты называют такую частоту раздражающих импульсов, при которой каждый последующий импульс совпадает с фазой повышенной возбудимости (рис. 4, A) и соответственно вызывает тетанус наибольшей амплитуды. Пессимумом частоты называют более высокую частоту раздражения, при которой каждый последующий импульс тока попадает в фазу рефрактерности (рис. 4, A), в результате чего амплитуда тетануса значительно уменьшается.
Работа скелетной мышцы
Сила сокращения скелетной мышцы определяется 2 факторами:
числом ДЕ, участвующих в сокращении;
частотой сокращения мышечных волокон.
Работа скелетной мышцы совершается за счет согласованного изменения тонуса (напряжения) и длины мышцы во время сокращения.
Виды работы скелетной мышцы:
динамическая преодолевающая работа совершается, когда мышца, сокращаясь, перемещает тело или его части в пространстве;
статическая (удерживающая) работа выполняется, если благодаря сокращению мышцы части тела сохраняются в определенном положении;
динамическая уступающая работа совершается, если мышца функционирует, но при этом растягивается, так как совершаемого ею усилия недостаточно, чтобы переместить или удержать части тела.
Во время выполнения работы мышца может сокращаться:
изотонически – мышца укорачивается при постоянном напряжении (внешней нагрузке); изотоническое сокращение воспроизводится только в эксперименте;
изометричеки – напряжение мышцы возрастает, а ее длина не изменяется; мышца сокращается изометрически при совершении статической работы;
ауксотонически – напряжение мышцы изменяется по мере ее укорочения; ауксотоническое сокращение выполняется при динамической преодолевающей работе.
Правило средних нагрузок – мышца может совершить максимальную работу при средних нагрузках.
Утомление – физиологическое состояние мышцы, которое развивается после совершения длительной работы и проявляется снижением амплитуды сокращений, удлинением латентного периода сокращения и фазы расслабления. Причинами утомления являются: истощение запаса АТФ, накопление в мышце продуктов метаболизма. Утомляемость мышцы при ритмической работе меньше, чем утомляемость синапсов. Поэтому при совершении организмоммышечной работыутомление первоначально развивается на уровне синапсов ЦНС и нейро-мышечных синапсов.
Структурная организация и сокращение гладких мышц
Структурная организация. Гладкая мышца состоит из одиночных клеток веретенообразной формы ( миоцитов ), которые располагаются в мышце более или менее хаотично. Сократительные филламенты расположены нерегулярно, вследствие чего отсутствует поперечная исчерченность мышцы.
Механизм сокращения аналогичен таковому в скелетной мышце, но скорость скольжения филламентов и скорость гидролиза АТФ в 100–1000 раз ниже, чем в скелетной мускулатуре.
Механизм сопряжения возбуждения и сокращения. При возбуждении клетки Cа++ поступает в цитоплазму миоцита не только из саркоплазматичекого ретикулума, но и из межклеточного пространства. Ионы Cа++ при участии белка кальмодулина активируют фермент (киназу миозина), который переносит фосфатную группу с АТФ на миозин. Головки фосфорилированного миозина приобретают способность присоединяться к актиновым филламентам.
Сокращение и расслабление гладких мышц. Скорость удаления ионов Са++ из саркоплазмы значительно меньше, чем в скелетной мышце, вследствие чего расслабление происходит очень медленно. Гладкие мышцы совершают длительные тонические сокращения и медленные ритмические движения. Вследствие невысокой интенсивности гидролиза АТФ гладкие мышцы оптимально приспособлены для длительного сокращения, не приводящего к утомлению и большим энергозатратам.
Физиологические свойства мышц
Общими физиологическими свойствами скелетных и гладких мышц являются возбудимость и сократимость. Сравнительная характеристика скелетных и гладких мышц приведена в табл. 6.1.Физиологические свойстваи особенности сердечной мускулатуры рассматриваются в разделе «Физиологические механизмы гомеостаза».
Таблица 7.1. Сравнительная характеристика скелетных и гладких мышц
Свойство | Скелетные мышцы | Гладкие мышцы |
Скорость деполяризации | медленная | |
Период рефрактерности | короткий | длительный |
Характер сокращения | быстрые фазические | медленные тонические |
Энергозатраты | ||
Пластичность | ||
Автоматия | ||
Проводимость | ||
Иннервация | мотонейронами соматической НС | постганглионарными нейронами вегетативной НС |
Осуществляемые движения | произвольные | непроизвольные |
Чувствительность к химическим веществам | ||
Способность к делению и дифференцировке |
Пластичность гладких мышц проявляется в том, что они могут сохранять постоянный тонус как в укороченном, так и в растянутом состоянии.
Проводимость гладкой мышечной ткани проявляется в том, что возбуждение распространяется от одного миоцита к другому через специализированные электропроводящие контакты (нексусы).
Свойство автоматии гладкой мускулатуры проявляется в том, что она может сокращаться без участиянервной системы, за счет того, что некоторые миоциты способны самопроизвольно генерировать ритмически повторяющиеся потенциалы действия.
Различают несколькоформ и типов мышечных сокращений.
1. Динамическаяформа мышечного сокращения. При такомтипе сокращений изменяется длина мышцы,но не изменяется напряжение. Эта формавключает два типа:
а) Изотоническийтип или концентрационный (мышцаукорачивается, но не изменяет своегонапряжения). Например, ходьба.
б) Эксцентрическийтип. Если нагрузка на мышцу больше, чемее напряжение, то мышца растягивается.Например, при опускании тяжелогопредмета.
2 Статическая формамышечного сокращения. Эта форманаблюдается при поддержании позы илипреодолении силы земного притяжения.
Данная формавключает один тип мышечного сокращения– изометрический. При изометрическомсокращении мышца изменяет свое напряжение,но не изменяет длины.
3. Форма ауксотоническогосокращения или смешанная.
Деление на формыи типы мышечных сокращений являетсяусловным т.к. все сокращения являютсясмешанными. Однако преобладает какой-тоодин тип.
Режимы сокращения мышц.
Характер или режимсокращения мышцы зависит от частотыимпульсов, которые поступают отмотонейрона.
Выделяют одиночныеи тетанические мышечные сокращения.
Если на мышцуподействовать одиночным импульсом, топроисходит одиночноемышечное сокращение, в котором выделяют несколько фаз:
1. Латентный(скрытый) период – время после действияраздражителя до начала сокращения.
2. Фаза укорочения(при изотоническом сокращении) или фазанапряжения (при изометрическомсокращении).
3. Фаза расслабления.
Одиночное мышечноесокращение характеризуется не значительнойутомляемостью, но при этом мышца неспособна реализовать свои возможности.
Тетаническоемышечное сокращение. Если на мышечное волокно воздействуютдва быстро следующих друг за другомимпульса, то сокращения накладываютсяи возникает сильное сокращение.
Наложение двухследующих друг за другом импульсовназывается суммацией.
Выделяют два видасуммации:
1. Если второйраздражитель поступает в момент, когдамышца начала расслабляться, то криваяимеет вершину отдельную от вершиныпервого сокращения. Этот вид суммацииназывается неполной.
2. Если второйраздражитель поступает в момент, когдасокращение мышцы еще не дошло до вершиныт.е. мышца не начала расслабляться, тооба сокращения сливаются в единое целое.Этот вид суммации называется полной.
Длительное исильное сокращения мышцы, под влияниемритма импульсов с последующим расслаблениемназывается тетанусом. У человека тетанусможно получить при частоте 50-70 имп/сек.
Выделяют два видатетануса:
1. Зубчатый. Возникаетпри малой частоте подачи импульсов (до150 имп/cек).
2. Гладкий. Возникаетпри высоком ритме подачи импульсов(более 150 имп/cек).
При этом различаютоптимальный и пессимальный ритмы работымышцы.
Так, если частотаподачи и сила импульсов вызываетмаксимальный сократительный эффект,то это оптимальный ритм работы.Оптимальный ритм работы формируетсячерез фазу экзальтации (т.е.супернормальности).
Если частота подачиимпульсов и сила раздражителя слишкомвелики, то это вызывает снижение силысокращения. Такой ритм называетсяпессимальным. Этот ритм работы мышцыформируется через фазу абсолютнойрефрактерности.
− В зависимости от условий, в которых происходит мышечное сокращение, различают два его основных типа - изотоническое и изометрическое. Сокращение мышцы, при котором ее волокна укорачиваются, но напряжение остается постоянным, называется изотоническим. Изометрическим является такое сокращение, при котором мышца укоротиться не может, если оба ее конца закреплены неподвижно. В этом случае по мере развития сократительного процесса напряжение возрастает, а длина мышечных волокон остается неизменной.
В натуральных двигательных актах сокращения мышц смешанные: даже поднимая постоянный груз, мышца не только укорачивается, но и изменяет свое напряжение вследствие реальной нагрузки. Такое сокращение называется ауксотоническим.
− В зависимости от частоты стимуляции выделяют одиночные и тетанические сокращения.
Одиночное сокращение (напряжение) возникает при действии на мышцу одиночного электрического или нервного импульса. Волна возбуждения возникает в месте приложения электродов для прямого раздражения мышцы или в области нервно-мышечного соединения и отсюда распространяется вдоль всего мышечного волокна. В изотоническом режиме одиночное сокращениеикроножной мышцылягушки начинается через короткий скрытый (латентный) период - до 0,01 с, далее следуют фаза подъема (фаза укорочения) - 0,05 с и фаза спада (фаза расслабления) - 0,05-0,06 с. Обычно мышца укорачивается на 5-10 % от исходной длины. Как известно, продолжительность волны возбуждения (ПД) мышечных волокон варьирует, составляя величину порядка 1-10 мс (с учетом замедления фазы реполяризации в ее конце). Таким образом, длительность одиночного сокращения мышечного волокна, наступающего вслед за его возбуждением, во много раз превышает продолжительность ПД.
Мышечное волокно реагирует на раздражение по правилу «все или ничего», т.е. отвечает на все надпороговые раздражения стандартным ПД и стандартным одиночным сокращением. Однако сокращение целой мышцы при ее прямом раздражении находится в большой зависимости от силы стимуляции. Это связано с различной возбудимостью мышечных волокон и разным расстоянием их от раздражающих электродов, что ведет к неодинаковому количеству активированных мышечных волокон. При пороговой силе стимула сокращение мышцы едва заметно, потому что в реакцию вовлекается лишь небольшое количество волокон. При увеличении силы раздражения число возбужденных волокон растет, пока все волокна не окажутся сокращенными, и тогда достигается максимальное сокращение мышцы. Дальнейшее усиление стимулов прироста амплитуды сокращения не вызывает.
В естественных условиях мышечные волокна работают в режиме одиночных сокращений только при относительно низкой частоте импульсации мотонейронов, когда интервалы между последовательными ПД мотонейронов превышают длительность одиночного сокращения иннервируемых ими мышечных волокон. Еще до прихода следующего импульса от мотонейронов мышечные волокна успевают полностью расслабиться. Новое сокращение возникает после полного расслабления мышечных волокон. Такой режим работы обусловливает незначительную утомляемость мышечных волокон. При этом ими развивается относительно небольшое напряжение.
Тетаническое сокращение - это длительное слитное сокращение скелетных мышц. В его основе лежит явление суммации одиночных мышечных сокращений. При нанесении на мышечное волокно или целую мышцу двух быстро следующих друг за другом раздражении возникающее сокращение будет иметь большую амплитуду. Сократительные эффекты, вызванные первым и вторым раздражениями, как бы складываются, происходит суммация, или суперпозиция, сокращений, поскольку нити актина и миозина дополнительно скользят друг относительно друга. При этом в сокращение могут вовлекаться ранее не сокращавшиеся мышечные волокна, если первый стимул вызвал у них подпороговую деполяризацию, а второй увеличивает ее до критической величины. При получении суммации в одиночном волокне важно, чтобы второе раздражение наносилось после исчезновения ПД, т.е. после рефрактерного периода. Естественно, что суперпозиция сокращений наблюдается и при стимуляции моторного нерва, когда интервал между раздражениями короче всей длительности сократительного ответа, в результате чего и происходит слияние сокращений.
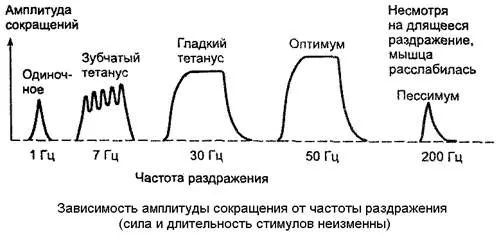
При сравнительно низких частотах наступает зубчатый тетанус, при большой частоте - гладкий тетанус (рис. 13).
Рис. 13. Сокращения икроножной мышцы лягушки при увеличении частоты раздражения седалищного нерва. Суперпозиция волн сокращения и образованиеразных видовтетануса.
а - одиночное сокращение (Г = 1 Гц); б, в - зубчатый тетанус (Г= 15-20 Гц); г, д - гладкий тетанус и оптимум (Г = 25-60 Гц); е - пессимум - расслабление мышцы во время раздражения (Г= 120 Гц).
Их амплитуда больше величины максимального одиночного сокращения. Напряжение, развиваемое мышечными волокнами при гладком тетанусе, обычно в 2-4 раза больше, чем при одиночном сокращении. Режим тетанического сокращения мышечных волокон в отличие от режима одиночных сокращений быстрее вызывает их утомление и поэтому не может поддерживаться длительное время. Из-за укорочения или полного отсутствия фазы расслабления мышечные волокна не успевают восстановить энергетические ресурсы, израсходованные в фазе укорочения. Сокращение мышечных волокон при тетаническом режиме с энергетической точки зрения происходит «в долг».
До сих пор нет общепризнанной теории, объясняющей, почему напряжение, развиваемое при тетанусе, или суперпозиции сокращений, гораздо больше, чем при одиночном сокращении. Во время кратковременной активации мышцы вначале одиночного сокращения в поперечных мостиках между нитями актина и миозина возникает упругое напряжение. Однако недавно было показано, что такой активации недостаточно для прикрепления всех мостиков. Когда она более длительная, обеспечиваемая ритмической стимуляцией (например, при тетанусе), их прикрепляется больше. Количество поперечных мостиков, связывающих актиновые и миозиновые филаменты (а, следовательно, и развиваемая мышцей сила), согласно теории скользящих нитей, зависит от степени перекрывания толстых и тонких нитей, а значит, и от длины саркомера или мышцы.
Высвобождение Са 2+ при тетанусе. Если стимулы поступают с высокой частотой (не менее 20 Гц), уровень Са 2 + в интервалах между ними остается высоким, потому что кальциевый насос не успевает вернуть все ионы в продольную систему саркоплазматического ретикулума. В таких условиях отдельные сокращения почти полностью сливаются. Это состояние устойчивого сокращения, или тетанус, наблюдается в том случае, когда промежутки между стимулами (или потенциалами действия в клеточной мембране) меньше примерно 1/3 длительности каждого из одиночных сокращений. Следовательно, частота стимуляции, необходимая для их слияния, тем ниже, чем больше их длительность; по этой причине она зависит от температуры. Минимальный промежуток времени между последовательными эффективными стимулами во время тетануса не может быть меньше рефрактерного периода, который приблизительно соответствует длительности потенциала действия.
Как выяснилось, амплитуда гладкого тетануса колеблется в широких пределах в зави- \ симости от частоты стимуляции нерва. При некоторой оптимальной (достаточно высокой) частоте стимуляции амплитуда гладкого тетануса становится наибольшей. Такой гладкий тетанус получил название оптимума. При дальнейшем повышении частоты стимуляции нерва развивается блок проведения возбуждения в нервно-мышечных синапсах, приводящий к расслаблению мышцы в ходе стимуляции нерва - пессимум Введенского. Частота стимуляции нерва, при которой наблюдается пессимум, получила название пессимальной (см. рис. 6.4).
В эксперименте легко обнаруживается, что уменьшенная в ходе пессимальной ритмической стимуляции нерва амплитуда мышечного сокращения моментально возрастает при возвращении частоты раздражения от пессимальной к оптимальной. В этом наблюдении - хорошее доказательство того, что пессимальное расслабление мышцы не является следствием утомления, истощения энергоемких соединений, а является следствием особых соотношений, складывающихся на уровне пост- и пресинаптических структур нервно-мышечного синапса. Пессимум Введенского можно получить и при прямом, но более частом раздражении мышцы (около 200 имп/с).
Контрактура. Контрактурой называется состояние обратимого местного устойчивого сокращения. Оно отличается от тетануса отсутствием распространяющегося потенциала действия. При этом может наблюдаться длительная локальная деполяризация мышечной мембраны, например при калиевой контрактуре, или же мембранный потенциал, близкий к уровню покоя, в частности при кофеиновой контрактуре. Кофеин при нефизиологически высоких (миллимолярных) концентрациях проникает в мышечные волокна и, не вызывая возбуждения мембраны, способствует высвобождению Са 2+ из саркоплазматического ретикулума; в результате развивается контрактура. При калиевой контрактуре степень стойкой деполяризации и сократительного напряжения волокна зависит от концентрации К + в наружном растворе.