Отличительным свойством гладкомышечных клеток является. Физиология гладких мышц
Нормальная физиология: конспект лекций Светлана Сергеевна Фирсова
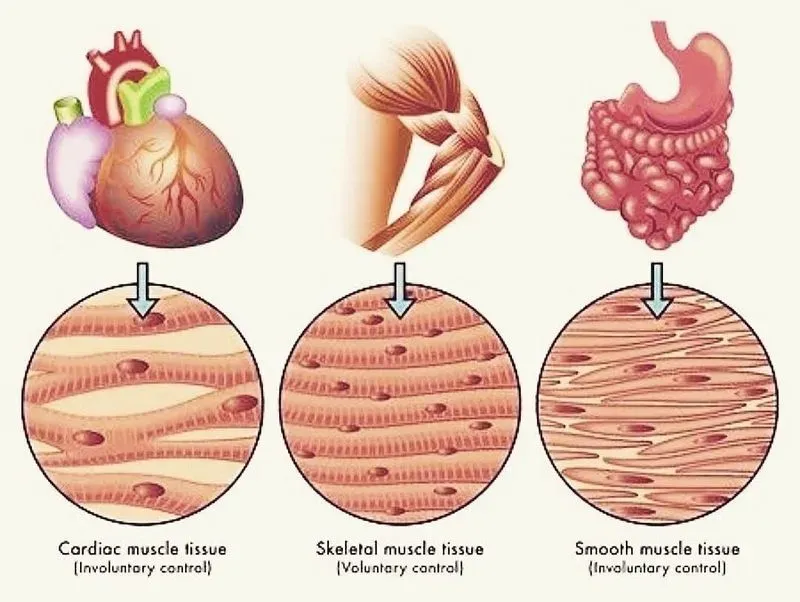
1. Физические и физиологические свойства скелетных, сердечной и гладких мышц
По морфологическим признакам выделяют три группы мышц:
1) поперечно-полосатые мышцы (скелетные мышцы);
2) гладкие мышцы;
3) сердечную мышцу (или миокард).
Функции поперечно-полосатых мышц:
1) двигательная (динамическая и статическая);
2) обеспечения дыхания;
3) мимическая;
4) рецепторная;
5) депонирующая;
6) терморегуляторная.
Функции гладких мышц:
1) поддержание давления в полых органах;
2) регуляция давления в кровеносных сосудах;
3) опорожнение полых органов и продвижение их содержимого.
Функция сердечной мышцы – насосная, обеспечение движения крови по сосудам.
Физиологические свойстваскелетных мышц:
1) возбудимость (ниже, чем в нервном волокне, что объясняется низкой величиной мембранного потенциала);
2) низкая проводимость, порядка 10–13 м/с;
3) рефрактерность (занимает по времени больший отрезок, чем у нервного волокна);
4) лабильность;
5) сократимость (способность укорачиваться или развивать напряжение).
Различают два вида сокращения:
а) изотоническое сокращение (изменяется длина, тонус не меняется);
б) изометрическое сокращение (изменяется тонус без изменения длины волокна). Различают одиночные и титанические сокращения. Одиночные сокращения возникают при действии одиночного раздражения, а титанические возникают в ответ на серию нервных импульсов;
6) эластичность (способность развивать напряжение при растягивании).
Физиологические особенности гладких мышц.
Гладкие мышцы имеют те же физиологические свойства, что и скелетные мышцы, но имеют и свои особенности:
1) нестабильный мембранный потенциал, который поддерживает мышцы в состоянии постоянного частичного сокращения – тонуса;
2) самопроизвольную автоматическую активность;
3) сокращение в ответ на растяжение;
4) пластичность (уменьшение растяжения при увеличении растяжения);
5) высокую чувствительность к химическим веществам.
Физиологической особенностью сердечной мышцы является ее автоматизм. Возбуждение возникает периодически под влиянием процессов, протекающих в самой мышце. Способностью к автоматизму обладают определенные атипические мышечные участки миокарда, бедные миофибриллами и богатые саркоплазмой.
автора Светлана Сергеевна ФирсоваИз книги Нормальная физиология: конспект лекций автора Светлана Сергеевна ФирсоваИз книги Нормальная физиология: конспект лекций автора Светлана Сергеевна ФирсоваИз книги Нормальная физиология: конспект лекций автора Светлана Сергеевна ФирсоваавтораИз книги Медицинская физика автора Вера Александровна Подколзинаавтора Марина Геннадиевна ДрангойИз книги Анализы. Полный справочник автора Михаил Борисович ИнгерлейбИз книги Нормальная физиология автора Николай Александрович АгаджанянИз книги Атлас: анатомия и физиология человека. Полное практическое пособие автора Елена Юрьевна ЗигаловаИз книги Полный справочник анализов и исследований в медицине автора Михаил Борисович ИнгерлейбИз книги Массаж при заболеваниях позвоночника автора Галина Анатольевна ГальперинаИз книги Гимнастика будущего автора Анатолий СительИз книги Справочник ветеринара. Руководство по оказанию неотложной помощи животным автора Александр ТалькоИз книги Скульптурная гимнастика для мышц, суставов ивнутренних органов. автора Анатолий СительИз книгиСкандинавская ходьба. Секреты известного тренера автора Анастасия ПолетаеваГладкие мышцы, формирующие стенки (мышечные слои) внутренних органов делят на два типа – висцеральные (т. е. внутренние) гладкие мышцы, выстилающие стенки желудочно-кишечного тракта и мочевыделительного тракта, и унитарные – гладкие мышцы, расположенные в стенках сосудов, в зрачке и хрусталике глаза и у корней волос кожного покрова (мышцы, взъерошивающие шерсть у животных). Эти мышцы построены из веретенообразных одноядерных клеток, не имеющих поперечной исчерченности, что обусловлено хаотичным расположением сократительных белков в их волокнах. Мышечные волокна относительно коротки (от 50 до 200 мкм), они имеют ветвления на обоих концах и плотно прилегают друг к другу, образуя длинные и тонкие цилиндрические пучки диаметром 0,05-0,01 мм, которые ветвятся и соединяются с другими пучками. Их сеть образует во внутренних органах либо слои (пласты), либо еще более толстые пучки.
Соседние клетки в гладких мышцах функционально связаны между собой низкоомными электрическими контактами – нексусами. За счет этих контактов потенциалы действия и медленные волны деполяризации беспрепятственно распространяются с одного мышечного волокна на другое. Поэтому несмотря на то, что двигательные нервные окончания расположены на небольшом числе мышечных волокон, в сократительную реакцию вовлекается вся мышца. Следовательно, гладкие мышцы представляют собой не только морфологический, но и функциональный синцитий.
Как и в скелетных мышцах, сократительные белки гладких мышц активируются в результате повышения концентрации ионов кальция в саркоплазме. Однако кальций поступает не из цистерн саркоплазматического ретикулума, как у скелетных мышц, а из внеклеточной среды, по градиенту концентрации, через плазматическую мембрану клетки, по медленным потенциалчувствительным кальциевым каналам, которые активируются в результате деполяризации мембраны при её возбуждении. Это существенно влияет на развитие потенциала действия гладкомышечных клеток, что наглядно отражает кривая ПД (Рис. 12. 1).
Рис.12. Потенциал действия (1) и кривая
сокращения (2) гладкомышечной клетки.
А – фаза деполяризации (Na + - вход);
Б – «кальциевое плато» (Ca 2+ - вход);
В – фаза реполяризации (К + - выход);
(пунктиром обозначен ПДскелетной мышцы)
Медленный, но достаточно существенный по величине входящий кальциевый ток формирует на кривой ПД характерное «кальциевое плато», которое не позволяет быстро деполяризовать мембрану, что приводит к значительному возрастанию продолжительности рефрактерного периода. Удаляется кальций из клетки еще медленнее, через Ca 2+ - АТФ-азы плазматической мембраны. Все это существенно сказывается как на характеристиках возбудимости, так и на сократительной способности гладких мышц. Гладкие мышцы гораздо менее возбудимы, чем поперечнополосатые и возбуждение по ним распространяется с очень небольшой скоростью – 2-15 см/с., кроме того, они сокращаются и расслабляются очень медленно, а время одиночного сокращения может продолжаться несколько секунд.
Из-за продолжительного рефрактерного периода длительность потенциала действия гладкомышечного волокна практически совпадает со временем поступления и удаления ионов кальция из клетки, то есть время развития ПД и длительность сокращения практически совпадают (Рис.12. 2) В результате гладкие мышцы практически не способны к формированию классического тетануса. Из-за очень медленного расслабления слияние одиночных сокращений («тетанус гладких мышц») возникает даже при низкой частоте раздражения и является, в большей степени, результатом медленного волнообразного вовлечения в длительное сокращение клеток, соседних с раздражаемой.
Гладкие мышцы способны к осуществлению относительно медленных и длительных тонических сокращений. Медленные, имеющие ритмический характер, сокращения гладких мышц желудка, кишечника, мочеточников и других органов обеспечивают перемещение содержимого этих органов. Длительные тонические сокращения гладких мышц особенно хорошо выражены в сфинктерах полых органов, которые препятствуют выходу содержимого этих органов.
Гладкие мышцы стенок кровеносных сосудов, особенно артерий и артериол, также находятся в состоянии постоянного тонического сокращения. Изменение тонуса мышц стенок артериальных сосудов влияет на величину их просвета и, следовательно, на уровень кровяного давления и кровоснабжения органов.
Важным свойством гладких мышц является их пластичность, т. е. способность сохранять приданную им при растяжении длину. Скелетная мышца в норме почти не обладает пластичностью. Эти различия хорошо наблюдать при медленном растяжении гладкой и скелетной мышцы. При удалении растягивающего груза скелетная мышца быстро укорачивается, а гладкая остается растянутой. Высокая пластичность гладких мышц имеетбольшое значениедля нормального функционирования полых органов. Благодаря высокой пластичности гладкая мышца может быть полностью расслаблена как в укороченном, так и в растянутом состоянии. Так, например, пластичность мышц мочевого пузыря по мере его наполнения предотвращает избыточное повышение давления внутри его.
Адекватным раздражителем для гладких мышц является их быстрое и сильное растяжение, что вызывает их сокращение, обусловленное нарастающей при растяжении деполяризацией клеток. Частота потенциалов действия (и соответственно частота сокращений.) тем больше, чем больше и быстрее растягивается гладкая мышца. Благодаря такому механизму обеспечивается, в частности, продвижение пищевого комка по пищеварительному тракту. Растянутая комком пищи мышечная стенка кишки отвечает сокращением и таким образом проталкивает комок в следующий участок кишки. Сокращение, индуцируемое растяжением, играетважную рольв авторегуляции тонуса кровеносных сосудов, а также обеспечивает непроизвольное (автоматическое) опорожнение переполненного мочевого пузыря в тех случаях, когда нервная регуляция отсутствует в результате повреждения спинного мозга.
Нервная регуляция гладких мышц осуществляется через симпатические и парасимпатические волокна вегетативнойнервной системы.
Особенность висцеральных гладкомышечных клеток состоит в том, что они способны сокращаться и при отсутствии прямых нервных влияний в условиях их изоляции и денервации, и даже после блокады нейронов вегетативных ганглиев.
В этом случае сокращения возникают не в результате передачи нервных импульсов с нерва, а вследствие активности собственных клеток ( пейсмекеров ), которые идентичны по структуре другим мышечным клеткам, но отличаются по электрофизиологическим свойствам – обладают автоматией. В этих клетках активность мембранных ионных каналов отрегулирована таким образом, что их мембранный потенциал не уравновешивается, а постоянно «дрейфует». В результате на мембране регулярно возникают препотенциалы или пейсмекерные потенциалы, с определенной частотой деполяризующие мембрану до критического уровня. При возникновении потенциала действия в пейсмекере возбуждение распространяется от них к соседним, что приводит к их возбуждению и сокращению. В результате последовательно сокращается один участок мышечного слоя за другим.
Из этого следует, что висцеральные гладкие мышцы контролируются вегетативной нервной системой, которая осуществляет в отношении этих мышц не пусковую, а настроечную, регулирующую, функцию. Это означает, что сама активность висцеральных гладких мышц возникает спонтанно, без нервных влияний, но уровень этой активности (сила и частота сокращений) изменяется под влиянием вегетативной нервной системы. В частности, изменяя скорость «дрейфа» мембранного потенциала, нервные импульсы вегетативных волокон воздействуют на частоту сокращений висцеральных гладкомышечных волокон.
Унитарные гладкие мышцы также могут быть спонтанно активными, но они сокращаются в основном под влиянием нервных импульсов вегетативных волокон. Их особенность состоит в том, что приходящий к ним одиночный нервный импульс не способен вызвать сокращение, в ответ возникает лишь временная подпороговая деполяризация мембраны мышечной клетки. Только когда по вегетативному нервному волокну следует серия импульсов с частотой около 1 импульса в 1 сек. и более, возможно развития потенциала действия мышечного волокна и его сокращение. То есть, унитарныемышечные волокна«суммируют» нервные импульсы и отвечают на раздражение когда частота импульсов достигает определенной величины.
В унитарной гладкой мышце, как и в висцеральных гладких мышцах, возбужденные мышечные клетки оказывают влияние на соседние клетки. В результате возбуждение захватывает много клеток (отсюда название этих мышц – унитарные, т. е. состоящие из унит – «единиц» с большим числом мышечных волокон в каждой из них).
В нервной регуляции сокращения гладких мышц участвуют два медиатора – ацетилхолин (АХ) и адреналин (норадреналин). Способ действия АХ в гладких мышцах такой же, как и в скелетных: АХ увеличивает ионную проницаемость мембраны, вызывая ее деполяризацию. Механизм действия адреналина неизвестен. Скелетные мышечные волокна реагируют на действие медиатора только в области концевой пластинки (нервно-мышечного синапса), тогда как гладкомышечные волокна отвечают на действие медиатора независимо от места его приложения. Поэтому на гладкие мышцы могут влиять медиаторы, содержащиеся в крови (например, адреналин, оказывающий длительное влияние на гладкие мышцы, вызывает их сокращение).
Из всего вышесказанного следует еще одна характерная особенность гладких мышц – их сокращение не требует больших энергетических затрат.
Гладкие мышцы образуют стенки (мышечный слой) внутренних органов и кровеносных сосудов. В миофибриллах гладких мышц нет поперечной исчерченности. Это обусловлено хаотичным расположением сократительных белков. Волокна гладких мышц относительно короче.
Гладкие мышцы менее возбудимы, чем поперечнополосатые. Возбуждение в гладких мышцах может передаваться с одного волокна на другое, в отличие от нервных волокон и волокон поперечнополосатых мышц.
Сокращение гладкой мускулатуры происходит более медленно и длительно.
Рефрактерный период в гладких мышцах более продолжителен, чем в скелетных.
Важным свойством гладкой мышцы является ее большая пластичность, т.е. способность сохранять приданную растяжением длину без изменения напряжения.
Особенностью гладких мышц является их способность к автоматической деятельности, которая обеспечивается нервными элементами, заложенными в стенках гладкомышечных органов.
Адекватным раздражителем для гладких мышц является их быстрое и сильное растяжение, что имеет большое значение для функционирования многих гладкомышечных органов (мочеточник, кишечник и другие полые органы)
Особенностью гладких мышц является также их высокая чувствительностьк некоторым биологически активным веществам (ацетилхолин, адреналин, норадреналин, серотонин и др.).
Гладкие мышцы иннервируются симпатическими и парасимпатическими вегетативными нервами, которые, как правило, оказывают противоположное влияние на их функциональное состояние.
Двигательные единицы, их классификация. Физические свойства мышц. Сила и работа мышц. Закон силы(ДОПИСАТЬ)
Каждое двигательное нервное волокно является отростком нервной клетки - мотонейрона, расположенного в переднем роге спинного мозга или в двигательном ядре черепного нерва. В мышце двигательное волокно ветвится и иннервирует не одно, а целую группу мышечных волокон. Мотонейрон вместе с группой иннервируемых им мышечных волокон называется двигательной единицей.
Мотонейроны бывают большие и малые.
Малые мотонейроны имеют тонкие аксоны и иннервируют небольшое количество (десятки) мышечных волокон, образуя малые двигательные единицы. Большие мотонейроны имеют толстые аксоны, которые иннервируют большое количество мышечных волокон (до нескольких тысяч), образуя большие двигательные единицы. Малые двигательные единицы входят в состав главным образом мелких мышц (пальцев рук, лица и др.), однако они входят также и в состав крупных мышц. Малые двигательные единицы обеспечивают быстрые и тонкие движения (например, движения пальцев рук). Большие двигательные единицы входят в состав преимущественно крупных мышц туловища и конечностей. Эти мышцы осуществляют относительно менее тонкие и более медленные движения, чем, например, движения пальцев рук. Малые мотонейроны (низкопороговые) возбуждаются легче и быстрее по сравнению с большими (высокопороговыми).
Гладкие мышцы построены из мышечных волокон, которые имеют диаметр от 2 до 5 мкм и длину лишь от 20 до 500 мкм, что значительно меньше, чем в скелетных мышцах, волокна которых имеют диаметр больше в 20 раз, а длину - в тысячи раз. Они не имеют поперечной исчерченности. Механизм сокращения гладких мышечных волокон принципиально такой же, как в лоперечнопосмугованих. Он построен на взаимодействии между сократительные белки актина и миозина, хотя существуют некоторые различия - для них не характерно упорядоченное расположение филаментов. Аналогом Z-линий в гладких мышцах является плотные тельца, которые содержатся в миоплазмы и соединены с клеточной мембраной и актиновыми филаментами. Сокращение различных гладких мышц длится от 0,2 с до 30 с. Абсолютная сила их составляет 4-6 кг / см2, в скелетных мышцах - 3-17 кг / см2.
Типы гладких мышц: гладкие мышцы разделяют на висцеральные, или унитарные, полиэлементных, или мультиунитарни, и гладкиемышцы сосудов, обладающие свойствами обоих предыдущих типов.
Висцеральные, или унитарные мышцы содержатся в стенках полых органов - пищеварительного канала, матки, мочеточников, желчного и мочевого пузыря. их особенностью является то, что они передают возбуждение от клетки к клетке щелевыми контактами низкого сопротивления, что позволяет мышцам реагировать как функциональный синцитий, то есть как одна клетка, отсюда и термин - унитарные мышцы. Они спонтанно активны, имеют водители ритма (пейсмекера), которые модулируются под влиянием гормонов или нейромедиаторов. Потенциал покоя для этих мышечных волокон не характерен, так как в активном состоянии клетки он низкий, во время ее торможения - высокий, а в состоянии покоя составляет около -55 мВ. Для них характерны так называемые синусоидальные медленные волны деполяризации, на которые накладываются пиковые ПД, продолжительностью от 10 до 50 мс (рис. 2.34).
Механизм генерации ПД гладких мышц и их сокращение в значительной степени инициируется ионами Са2 Сокращение возникает через 100-200 мс после возбуждения, а максимальное - развивается только через 500 мс после начала пика. Следовательно, сокращение гладких мышц является медленным процессом. Однаковисцеральные мышцыобладают высокой степенью электрического сопряжения между клетками, обеспечивает высокую координацию их сокращения.
Полиэлементных, или мультиунитарни гладкие мышцы состоят из отдельных единиц без соединительных мостиков, и ответ целого мышцы на раздражение состоит из ответа отдельных мышечных волокон. Каждое мышечное волокно иннервируется одним нервным окончанием, как в скелетных мышцах. К ним относятся мышцы радужки глаза, цилиарная мышцу глаза, пилоеректорни мышцы волос кожи. Они не имеют произвольной регуляции, сокращаются благодаря нервным импульсам, которые передаются через нервно-мышечные синапсы вегетативной нервной системы, нейромедиаторы которой могут вызвать как возбуждение, так и торможение.
Механизмы сокращения и расслабления гладких мышц
Механизм сопряжения возбуждения и сокращения отличается от подобного процесса, происходящего в скелетных мышцах, так как гладкие мышцы не содержат тропонина.
Последовательность процессов в гладких мышцах, что приводит к сокращению и расслаблению, имеет такие шаги:
1. При деполяризации клеточной мембраны открываются потенциалозалежни кальциевые каналы и ионы
РИС. 2.34.
Са 2+ входят в клетку с электрохимическим градиентом, концентрация ионов Са 2+ в клетке увеличивается.
2. Вход ионов Са 2+ через клеточную мембрану может вызвать дополнительный выход ионов Са 2+ с саркоплазматического ретикулума (СПР) через Са 2+ зависимые ворота кальциевых каналов. Гормоны и нейромедиаторы также стимулируют выход ионов Са 2+ с СПР через инозитолтрифосфатид (И-С-Ф) зависимые ворота кальциевых каналов.
3. внутриклеточной концентрации ионов Са 2+ увеличивается.
4. Ионы Са 2+ связываются с кальмодулином, регуляторным белком, который имеет 4 связывания Са 2+ и играет важную роль в активации ферментов. Кальций кальмодулиновий комплекс активирует фермент киназу легкой цепи миозина, что приводит к фосфорилирования молекул головки миозина. Миозин гидролизует АТФ, генерируется энергия и начинается цикл образования поперечных актино-миозиновых мостиков, скольжения актина по миозинових цепях. Фосфорилированные миозиновые мостики повторяют свой цикл, пока не дефосфорилюються миозинфосфатазою.
5. Дефосфорилирование миозина приводит к расслаблению мышечного волокна, или состояния остаточного напряжения благодаря образованным поперечным мостикам, пока не произойдет окончательная диссоциация кальций-кальмодулинового комплекса.
В процессе онтогенеза изменяются свойства возбудимых структур в связи с развитием опорно-двигательной системы и ее регуляцией.
Увеличивается масса мышц - от 23,3% массы тела у новорожденного до 44,2% в возрасте 17-18 лет. Растетмышечная тканьблагодаря удлинению и утолщению мышечных волокон, а не увеличению их количества.
У новорожденного ребенка активность натрий-калиевых насосов, расположенных в мембранах миоцитов, еще мала и поэтому концентрация ионов К + в клетке почти вдвое меньше, чем у взрослого человека, и только в 3 месяца начинает увеличиваться. ПД после рождения уже генерируются, однако имеют меньшую амплитуду и большую продолжительность. Генерация ПД мышечных волокон у новорожденных не блокируется тетродотоксином.
После рождения в нервных волокнах увеличивается длина и диаметр осевых цилиндров от 1-3 мкм до 7 мкм в 4 года, и завершается их формирования в 5-9 лет. До 9 лет заканчивается миелинизация нервных волокон. Скорость проведения возбуждения после рождения не превышает 50% скорости у взрослых и увеличивается в течение 5 лет. Рост скорости проведения обусловлено: увеличением диаметра нервных волокон, их миелинизацией, образованием ионных каналов и повышением амплитуды ПД. Уменьшение продолжительности ПД и соответственно фазы абсолютной рефрактерности приводит к увеличению количества ПД, которые может генерировать нервное волокно.
Рецепторный аппарат мышц развивается быстрее, чем формируются двигательные нервные окончания. Продолжительность нервно-мышечной передачи после рождения - 4,5 мс, у взрослого - 0,5 мс. В процессе онтогенеза возрастает синтез ацетилхолина, ацетилхолинэстеразы, плотность холинорецепторов конечной пластинки.
В процессе старения продолжительность ПД в возбудимых структурах увеличивается, а количество ПД, которые генерируют мышечные волокна в единицу времени (лабильность), уменьшается. Масса мышц уменьшается в связи с понижением интенсивности метаболизма.
Электрическая активность. Висцеральные гладкие мышцы характеризуются нестабильным мембранным потенциалом. Колебания мембранного потенциала независимо от нервных влияний вызывают нерегулярные сокращения, которые поддерживают мышцу в состоянии постоянного частичного сокращения - тонуса. Тонус гладких мышц отчетливо выражен в сфинктерах полых органов: желчном, мочевом пузырях, в месте перехода желудка в двенадцатиперстную кишку и тонкой кишки в толстую, а также в гладких мышцах мелких артерий и артериол.
В некоторых гладких мышцах, например мочеточника, желудка, лимфатических сосудов, ПД имеют продолжительное плато во время реполяризации. Платообразные ПД обеспечивают поступление в цитоплазму миоцитов значительного количества внеклеточного кальция, участвующего в последующем в активации сократительных белков гладкомышечных клеток. Ионная природа ПД гладкой мышцы определяется особенностями каналов мембраны гладкой мышечной клетки. Основную роль в механизме возникновения ПД играют ионы Са2+. Кальциевые каналы мембраны гладких мышечных клеток пропускают не только ионы Са2+, но и другие двухзарядные ионы (Bа 2+, Mg2+), а также Na+. Вход Са2+ в клетку во время ПД необходим для поддержания тонуса и развития сокращения, поэтому блокирование кальциевых каналов мембраны гладких мышц, приводящее к ограничению поступления иона Са2+ в цитоплазму миоцитов внутренних органов и сосудов, широко используется в практической медицине для коррекции моторики пищеварительного тракта и тонуса сосудов при лечении больных гипертонической болезнью.
Автоматия. ПД гладких мышечных клеток имеют авторитмический (пейсмекерный) характер, подобно потенциалам проводящей системы сердца. Пейсмекерные потенциалы регистрируются в различных участках гладкой мышцы. Это свидетельствует о том, что любые клетки висцеральных гладких мышц способны к самопроизвольной автоматической активности. Автоматия гладких мышц, т.е. способность к автоматической (спонтанной) деятельности, присуща многим внутренним органам и сосудам.
Реакция на растяжение. В ответ на растяжение гладкая мышца сокращается. Это вызвано тем, что растяжение уменьшает мембранный потенциал клеток, увеличивает частоту ПД и в конечном итоге - тонус гладкой мускулатуры. В организме человека это свойство гладкой мускулатуры служит одним из способов регуляции двигательной деятельности внутренних органов. Например, при наполнении желудка происходит растяжение его стенки. Увеличение тонуса стенки желудка в ответ на его растяжение способствует сохранению объема органа и лучшему контакту его стенок с поступившей пищей. Др. пр., растяжение мускулатуры матки растущим плодом служит одной из причин начала родовой деятельности.
Пластичность. Если растянуть висцеральную гладкую мышцу, то ее напряжение будет увеличиваться, однако если мышцу удерживать в состоянии удлинения, вызванным растяжением, то напряжение будет постепенно уменьшаться, иногда не только до уровня, существовавшего до растяжения, но и ниже этого уровня. Пластичность гладкой мускулатуры способствует нормальному функционированию внутренних полых органов.
Связь возбуждения с сокращением. В условиях относительного покоя можно зарегистрировать одиночный ПД. В основе сокращения гладкой мышцы, как и в скелетной, лежит скольжение актина по отношению к миозину, где ион Са2+ выполняет триггерную функцию.
В механизме сокращения гладкой мышцы имеется особенность, отличающая его от механизма сокращения скелетной мышцы. Эта особенность заключается в том, что прежде чем миозин гладкой мышцы сможет проявлять свою АТФазную активность, он должен быть фосфорилирован. Механизм фосфорилирования миозина гладкой мышцы осуществляется следующим образом: ион Са2+ соединяется с кальмодулином (кальмодулин - рецептивный белок для иона Са2+). Возникающий комплекс активирует фермент - киназу легкой цепи миозина, который в свою очередь катализирует процесс фосфорилирования миозина. Затем происходит скольжение актина по отношению к миозину, составляющее основу сокращения. Т.о. пусковым моментом для сокращения гладкой мышцы является присоединение иона Са2+ к кальмодулину, в то время как в скелетной и сердечной мышце пусковым моментом является присоединение Са2+ к тропонину.
Химическая чувствительность. Гладкие мышцы обладают высокой чувствительностью к различным физиологически активным веществам: адреналину, норадреналину, АХ, гистамину и др. Это обусловлено наличием специфических рецепторов мембраны гладкомышечных клеток.
Норадреналин действует на α- и β-адренорецепторы мембраны гладкомышечных клеток. Взаимодействие норадреналина с β-рецепторами уменьшает тонус мышцы в результате активации аденилатциклазы и образования циклического АМФ и последующего увеличения связывания внутриклеточного Са2+. Воздействие норадреналина на α-рецепторы тормозит сокращение за счет увеличения выхода ионов Са2+ из мышечных клеток.
АХ оказывает на мембранный потенциал и сокращение гладкой мускулатуры кишечника действие, противоположное действию норадреналина. Добавление АХ к препарату гладкой мышцы кишечника уменьшает мембранный потенциал и увеличивает частоту спонтанных ПД. В результате увеличивается тонус и возрастает частота ритмических сокращений, т. е. наблюдается тот же эффект, что и при возбуждении парасимпатических нервов. АХ деполяризует мембрану, увеличивает ее проницаемость для Na+ и Са++.
Похожая информация.